근막,fascia
- 광범위하고 비교가 어렵다.
- 서로 다른 조직을 연결: 해부 중에 개별 근육을 분리하는 것이 어려울 수 있습니다.
- 치밀 및 유륜 결합 조직, 심부 및 표면 근막, 골간막 및 격막, 외막, 골주위, 근내막, 골막, 신경혈관, 근육외 건막 포함
- 광범위한 조직화로 인해 근막은 구조 전체에 응력 및 하중을 분산시킬 수 있는 잠재력을 가지고 있습니다.
기계적으로 늘어나면 내부 힘 (internal force)이 발생합니다.
근육 사이 또는 조직(근육과 신경혈관) 사이의 근막은 힘 전달 경로를 나타낼 수 있습니다.
해부학적으로는 서로 다른 위치의 근육(대흉근, 상완이두근, FCR, 굴근지지대, 장장근) 근육 사이의 근막의 연속성을 보여줍니다.
힘이 전달되는 경로
- 근섬유(muscle fiber) - 신경근 접합부 (neuromusular junction)- 힘줄(tendon)-뼈 (bone)

근육 힘의 신경 조절은 근막 상호 작용에 의해 영향을 받는다.
근육 적응 메커니즘과 병태생리학을 재검토해야 할 것입니다.
근육 모델링은 근막 상호작용을 고려해야 합니다.
근육은 독립적인 작동자(actuator) 인가?
- 쥐 모델을 이용한 실험
전방 두개골 구획: 전경골근(TA), 긴무지신근(EHL) 및 긴수지신근(EDL)
EDL의 근위 및 원위 힘줄을 평가할 수 있습니다. 대퇴골에 있는 쥐의 EDL 근위부 부착
근육 배 근처와 엉덩이 구획 내의 결합 조직은 보존되었습니다
.
1. 근위 힘줄에서 측정된 힘이 항상 원위 힘줄의 힘과 동일하지는 않습니다.
EDL의 근위 및 원위 힘(능동 및 수동)은 다를 수 있습니다.
이러한 차이는 근육 사이의 근막이 절단되고 앞쪽 구획의 다른 근육이 제거될 때에도 지속됩니다.
결합 조직이 손상되지 않은 경우 근위 및 원위 근육(EDL) 힘줄의 능동 및 수동 장력이 다릅니다.
힘의 차이는 근육 간 및 근육 외 힘 전달을 나타냅니다.
2. 한 근육의 길이 변화는 다른 근육의 힘-길이 관계를 변화시킵니다.
TA + EHL 복합체의 길이와 힘 변화는 EDL 근육으로부터 힘 전달에 영향을 미칩니다
EDL의 힘-길이 관계는 직접적인 환경의 근육 및 결합 조직 구조의 특성에 따라 달라집니다.
일정하게 낮은 길이로 유지되는 근육(EDL)
근위 및 원위 EDL 힘줄의 활성 장력은 TA+ EHL의 함수에 따라 다릅니다.
***는 독립적인 구성요소로 작용하는 형태학적으로 정의된 근육의 개념이 근육에서 뼈로의 힘 전달을 설명하는 데 부적절할 수 있음을 시사합니다.
- 고양이 모델 실험
(근육의 생리학적 움직임에 따른 근육간 힘 전달 평가)
(무릎 각도를 조작하면 비복근과 족저근이 변경됨) 길이는 있지만 가자미근은 변경되지 않음
무릎 발목 각도 상수)
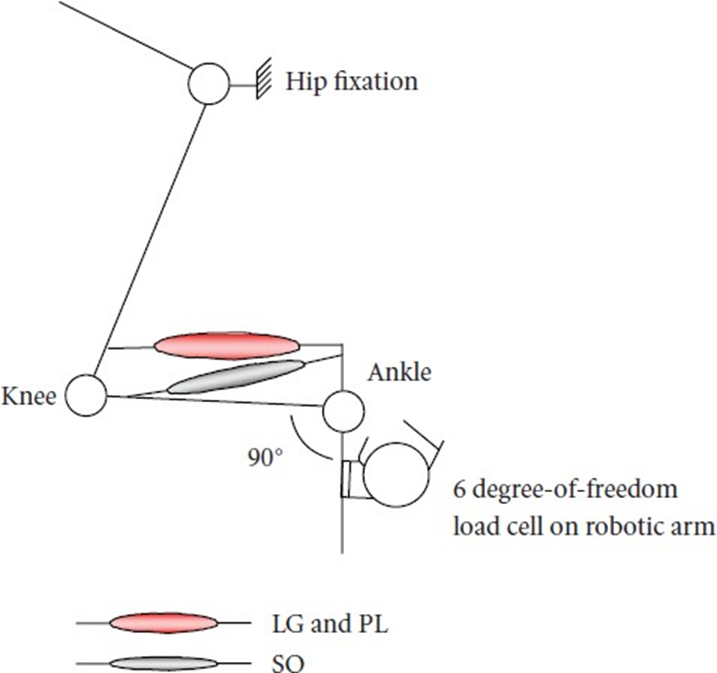
(무릎 각도를 70도에서 150도로 변경함 (그에 따라 위장 근육의 길이도 변경됨) 소레우스의 선택적 활성화에 의한 각도 토크에 영향을 주지 않음)
(상대적인 근육 운동은 반드시 이러한 근육 사이의 힘 전달을 의미하지 않음)
(따라서 위장 근육과 소레우스 간의 선형 힘 합은 존재함)
- 비선형성은 생리적 조건 하에서는 작게 나타났
- 근육을 독립적인 구동기로 모델링하는 것은 합리적으로 보였다.
- 잠재적 설명: 생리적 움직임 중 발가락 영역에서 인대의 변형이 발생할 수 있음 (생리학적 변위 중에만 선형 부분이 달성됨)
근육 작동
동작 유형에 따라 슬라이딩 필라멘트 및 크로스 브리지 이론이 잘 설명됨
길이와 힘의 변화에 대한 근이힘-길이 관계의 역학
- 역학에 대한 힘 및 에너지 요구는 대부분 과대평가되었다고 전해진다.
- A.F. Huxley (1980): 기대했던 바(이완 수축 분야) '몇 가지 놀라움을 경험할 것'. '특별한 특징이 진화하여 근육을 손상시키지 않고 과도하게 늘리는 것을 가능하게 한 것'을 상상한다고 했다.
크로스브릿지 모델은 힘 증가를 설명할 수 있다.
- 부착된 크로스브릿지의 비율이 증가함
- 크로스브릿지의 분포 거리가 증가함
- 그러나 느리게 늘리는 속도에 대한 힘 증가와 에너지 요구는 과대평가됨
- 모델의 개선이 필요함
이완 동작
- 잔여 힘 증
- 이전의 이완 동작 없이 수행된 등속성 동작과 비교하여 이완 동작 후에 장기간 지속되는 등속성 힘의 증가
- 힘 증가는 이완 동작 중 및 후에 관찰됨
- 힘 증가:
동안 (FE) 및 근육 작용 후 (PFE)
늘리기 크기에 따라 증가함
최대 50%까지 P0를 초과함
상승 및 하강 선에서 발생함
- 수동 힘 증가
근육 비활성 후에도 힘 증가가 존재함
근사수준에서 존재함 (사르코머의 특성을 나타냄)
등속성 동작에 앞서면 감소함 (이전 단축의 영향에 대한 시간 경과 약 1초)
티틴이 제거되면 사라짐
- 스프링 강성 증가(stiffness)
활성화되면, 티틴이 CA에 결합함
피엘리아 및 PEVK 도메인의 글루타메이트 풍부한 영역에 결합함
두 경우 모두 내재적 티틴의 강성이 증가함
- 따라서 특정 근육의 늘리기 크기에 대해, 근육이 개발하는 힘의 변화가 더 크게 됨
티틴의 근원적 영역이 알란 피라미드 근육필라멘트에 결합함
생체 내에서: 티틴의 N2A 영역이 액틴에 결합함
vitro에서: 티틴의 PEVK 도메인이 액틴에 결합함
* 증가된 수동적 강성 - CA가 액틴 및 티틴에 결합함에 의해
CA, 액틴 및 티틴 간의 결합 변화가 크로스브릿지 관계를 변경하지 않음
'생체역학(biomechanic)' 카테고리의 다른 글
| [생체 역학] 요추와 요통2 (0) | 2024.05.17 |
|---|---|
| [생체 역학] 관절의 기하학적 구조2 (0) | 2024.05.16 |
| [운동 역학] 근육의 힘과 속도의 관계 (0) | 2024.05.11 |
| [생체 역학] 요추와 요통 (0) | 2024.05.09 |
| [운동 역학] 근육의 힘과 길이의 관계 (0) | 2024.05.08 |




댓글